Human CCL5/RANTES ELISA Kit
 Protokollhäfte
Protokollhäfte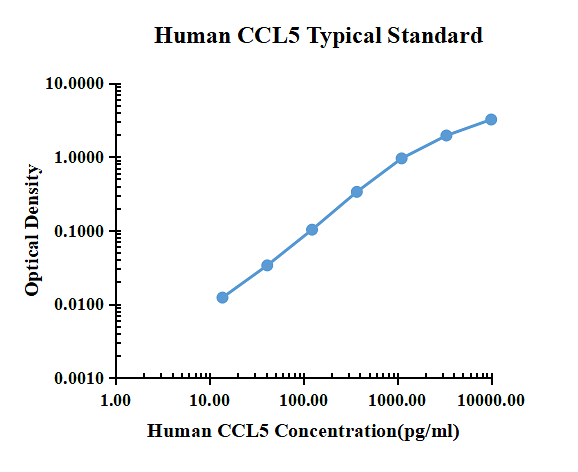
Nyckelfunktioner och detaljer
- Specifikation:
- Känslighet:
- Standardkurvområde:
- Standardkurvgradient:
- Antal inkubationer:
- Provvolym:
- Analystyp:
- Driftslängd:
-
Märke:
KAT.NR. : AEH0027
US$ Välj
US$ Välj
Storlek:
Trail, bulkstorlek eller anpassade önskemål Vänligen kontakta oss
 Produktinformation
Produktinformation
Bakgrund
RANTES (reglerad vid aktivering, normal T-cell uttryckt och förmodligen utsöndrad), även känd som CCL5, är en medlem av "CC"-underfamiljen av kemokiner. Det spelar en primär roll i det inflammatoriska immunsvaret via dess förmåga att kemoatrahera leukocyter och modulera deras funktion. cDNA för RANTES upptäcktes initialt genom subtraktiv hybridisering som en T-cellsspecifik sekvens. Humant RANTES-cDNA kodar för en mycket basisk 91 aminosyror (aa) prekursorpolypeptid med en 23 aa hydrofob signalpeptid som klyvs för att generera det 68 aa mogna proteinet. Human RANTES uppvisar ungefär 85 % homologi med mus RANTES på den härledda aa-nivån.
RANTES är en potent kemoattraherande medel för ett antal olika celltyper inklusive ostimulerade CD4+/CD45RO+ minnes-T-celler och stimulerade CD4+ och CD8+ T-celler med naiva och minnesfenotyper, NK-celler, basofiler, eosinofiler, dendritiska celler, mastceller, monocyter och mikroglia. Förutom dess effekter på migration kan RANTES aktivera ett antal celltyper inklusive T-celler, monocyter, neutrofiler, NK-celler, dendritiska celler och astrocyter. T-cellsaktivering kräver i allmänhet relativt höga RANTES-koncentrationer (~ 1 μM) och är beroende av aggregering av molekylen och association med cellyteglykosaminoglykaner (GAG). Huruvida detta [saknas några ord, förmodligen 'aktivitet sker in vivo'] är fortfarande oklart, även om intraperitonealt injicerade RANTES-mutanter som inte kan aggregera och/eller binda GAG hos möss inte kan attrahera leukocyter jämfört med kontroller av vildtyp. Andra in vivo-studier visar att RANTES knockout-möss uppvisar bristfällig rekrytering av leukocyter till platser med akut inflammation. - RANTES, är känd för att interagera med fyra identifierade sju transmembrana G-proteinkopplade receptorer: CCR1, CCR3, CCR4 och CCR5 (22-25). RANTES-stimulering kan initiera en mängd olika signalkaskader som är cellkontextberoende. Till exempel, i T-celler, kan RANTES stimulera förhöjningar av intracellulär Ca2+ och aktivering av fokal adhesion kinas (FAK), proteinkinas A, PI3-kinas, Rho GTPase och JAK/STAT signalvägar. Cytomegalovirusproteinet US28 uppvisar signifikant homologi med CC-kemokinreceptorer och har förmåga att binda RANTES. Membran-som spänner över US28 kan, beroende på sammanhanget, signalera på ett konstitutivt sätt, binda RANTES och initiera G-protein-förmedlade signalkaskader, eller binda RANTES och potentiellt förändra inflammatoriska svar.
RANTES-receptorn CCR5 är också den primära co-receptorn för R5 (M-tropic) varianter av HIV-1. Det har visat sig att RANTES, liksom de andra CCR5-liganderna, makrofaginflammatoriskt protein (MIP)-1 alfa och MIP-1 beta, kan kompetitivt hämma CCR5/HIV-1-interaktion och undertrycka virusinfektion in vitro. Dessa effekter kräver uppenbarligen inte helt intakt signalering från CCR5-receptorn. Följaktligen visar modifierade former av RANTES och icke-peptidföreningar som blockerar interaktionen mellan HIV-1 och CCR5 lovande för framtida terapier. Däremot visar flera rapporter att RANTES kan förbättra in vitro-replikation av X4 (T-tropic) varianter av HIV-1 som använder CXCR4 som en co-receptor snarare än CCR5. Denna aktivitet kräver vanligtvis relativt höga RANTES-koncentrationer (~μM) och är beroende av interaktion med cellyte-GAG, oligomerisering och aktivering av tyrosinkinas- och MAP-kinassignaleringskaskader.
RANTES är en potent kemoattraherande medel för ett antal olika celltyper inklusive ostimulerade CD4+/CD45RO+ minnes-T-celler och stimulerade CD4+ och CD8+ T-celler med naiva och minnesfenotyper, NK-celler, basofiler, eosinofiler, dendritiska celler, mastceller, monocyter och mikroglia. Förutom dess effekter på migration kan RANTES aktivera ett antal celltyper inklusive T-celler, monocyter, neutrofiler, NK-celler, dendritiska celler och astrocyter. T-cellsaktivering kräver i allmänhet relativt höga RANTES-koncentrationer (~ 1 μM) och är beroende av aggregering av molekylen och association med cellyteglykosaminoglykaner (GAG). Huruvida detta [saknas några ord, förmodligen 'aktivitet sker in vivo'] är fortfarande oklart, även om intraperitonealt injicerade RANTES-mutanter som inte kan aggregera och/eller binda GAG hos möss inte kan attrahera leukocyter jämfört med kontroller av vildtyp. Andra in vivo-studier visar att RANTES knockout-möss uppvisar bristfällig rekrytering av leukocyter till platser med akut inflammation. - RANTES, är känd för att interagera med fyra identifierade sju transmembrana G-proteinkopplade receptorer: CCR1, CCR3, CCR4 och CCR5 (22-25). RANTES-stimulering kan initiera en mängd olika signalkaskader som är cellkontextberoende. Till exempel, i T-celler, kan RANTES stimulera förhöjningar av intracellulär Ca2+ och aktivering av fokal adhesion kinas (FAK), proteinkinas A, PI3-kinas, Rho GTPase och JAK/STAT signalvägar. Cytomegalovirusproteinet US28 uppvisar signifikant homologi med CC-kemokinreceptorer och har förmåga att binda RANTES. Membran-som spänner över US28 kan, beroende på sammanhanget, signalera på ett konstitutivt sätt, binda RANTES och initiera G-protein-förmedlade signalkaskader, eller binda RANTES och potentiellt förändra inflammatoriska svar.
RANTES-receptorn CCR5 är också den primära co-receptorn för R5 (M-tropic) varianter av HIV-1. Det har visat sig att RANTES, liksom de andra CCR5-liganderna, makrofaginflammatoriskt protein (MIP)-1 alfa och MIP-1 beta, kan kompetitivt hämma CCR5/HIV-1-interaktion och undertrycka virusinfektion in vitro. Dessa effekter kräver uppenbarligen inte helt intakt signalering från CCR5-receptorn. Följaktligen visar modifierade former av RANTES och icke-peptidföreningar som blockerar interaktionen mellan HIV-1 och CCR5 lovande för framtida terapier. Däremot visar flera rapporter att RANTES kan förbättra in vitro-replikation av X4 (T-tropic) varianter av HIV-1 som använder CXCR4 som en co-receptor snarare än CCR5. Denna aktivitet kräver vanligtvis relativt höga RANTES-koncentrationer (~μM) och är beroende av interaktion med cellyte-GAG, oligomerisering och aktivering av tyrosinkinas- och MAP-kinassignaleringskaskader.
Typiska data
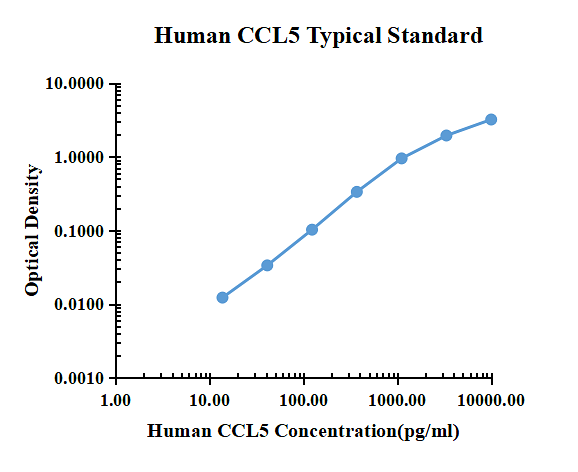
| pg/ml | O.D. | Genomsnittlig | Rättad | |
| 0.00 | 0.0151 | 0.0162 | 0.0157 | |
| 13.72 | 0.0271 | 0.0288 | 0.0280 | 0.0123 |
| 41.15 | 0.0489 | 0.0496 | 0.0493 | 0.0336 |
| 123.46 | 0.1175 | 0.1189 | 0.1182 | 0.1025 |
| 370.37 | 0.3486 | 0.3520 | 0.3503 | 0.3346 |
| 1111.11 | 1.0108 | 0.9250 | 0.9679 | 0.9522 |
| 3333.33 | 1.8759 | 2.0533 | 1.9646 | 1.9489 |
| 10000.00 | 3.2985 | 3.1585 | 3.2285 | 3.2128 |
Precision
| Intra-assay Precision | Inter-assay Precision | |||||
| Provnummer | S1 | S2 | S3 | S1 | S2 | S3 |
| 22 | 22 | 22 | 6 | 6 | 6 | |
| Genomsnitt (pg/ml) | 426.6 | 1631.6 | 3396 | 702.2 | 3412 | 8842.1 |
| Standardavvikelse | 37.6 | 128.2 | 288.6 | 44.6 | 194.4 | 506.5 |
| Variationskoefficient (%) | 8.8 | 7.9 | 8.5 | 6.4 | 5.7 | 5.7 |
Intra-assayprecision (precision inom en analys) Tre prover med känd koncentration testades tjugo gånger på en platta för att bedöma intra-assayprecisionen.
Inter-assayprecision (precision mellan analyser) Tre prover med känd koncentration testades sex gånger på en platta för att bedöma intra-assayprecisionen.
Spike Recovery
Spike-återvinningen utvärderades genom att spika 3 nivåer av humant CCL5 i ett hälsoprov från humant serum. Det otillsatta serumet användes som blank i detta experiment.
Återvinningen varierade från 90 % till 106 % med en total genomsnittlig återhämtning på 97 %.
Återvinningen varierade från 90 % till 106 % med en total genomsnittlig återhämtning på 97 %.
Exempelvärden
| Provmatris | Prov utvärderat | Område (pg/ml) | Detekterbar (%) | Medelvärde för detekterbar (pg/ml) |
| Serum | 30 | 943.0-38560.8 | 100 | 21207.7 |
Serum/plasma – Trettio prover från till synes friska frivilliga utvärderades med avseende på närvaron av CCL5 i denna analys. Ingen medicinsk historia var tillgänglig för donatorerna.
Nya produkter