Kit ELISA per IL-1β supersensibile per topi
Caratteristiche principali e dettagli
- Specifica:
- Sensibilità:
- Intervallo di curve standard:
- Gradiente curva standard:
- Numero di incubazioni:
- Volume del campione:
- Tipo di analisi:
- Durata dell'operazione:
-
Marca:
N. CAT. : AEMSS0029
US$ Scegli
US$ Scegli
Dimensioni:
Trail, dimensioni sfuse o richieste personalizzate Vi preghiamo di contattarci
 Dettagli del prodotto
Dettagli del prodotto
Sfondo
La famiglia di proteine dell'interleuchina 1 (IL-1) è costituita da IL-1 alfa, IL-1 beta e dall'antagonista del recettore IL-1 (IL-1ra). IL-1 alfa e IL-1 beta si legano agli stessi recettori sulla superficie cellulare e condividono funzioni biologiche. IL-1 non è prodotto da cellule non stimolate di individui sani, ad eccezione dei cheratinociti della pelle, di alcune cellule epiteliali e di alcune cellule del sistema nervoso centrale. Tuttavia, in risposta ad agenti infiammatori, infezioni o endotossine microbiche, si osserva un drammatico aumento della produzione di IL-1 da parte dei macrofagi e di vari altri tipi di cellule. IL-1 beta svolge un ruolo centrale nelle risposte immunitarie e infiammatorie, nel rimodellamento osseo, nella febbre, nel metabolismo dei carboidrati e nella fisiologia del GH/IGF-I. La produzione inappropriata o prolungata di IL-1 è stata implicata in una varietà di condizioni patologiche tra cui sepsi, artrite reumatoide, malattia infiammatoria intestinale, leucemia mieloide acuta e cronica, diabete mellito insulino-dipendente, aterosclerosi, danno neuronale e malattie legate all'invecchiamento.
IL-1 alfa e IL-1 beta sono polipeptidi strutturalmente correlati che mostrano circa il 25% di omologia a livello di amminoacidi (aa). Entrambi sono sintetizzati come precursori da 31 kDa che vengono successivamente scissi in proteine mature da circa 17,5 kDa. La scissione del precursore beta di IL-1 da parte di Caspase-1/ICE è un passaggio fondamentale nella risposta infiammatoria. Né IL-1 alfa né IL-1 beta contengono un tipico peptide segnale idrofobico, ma l'evidenza suggerisce che questi fattori possono essere secreti attraverso vie non-classiche. Una porzione di IL-1 alfa non trasformata può essere presente sulla membrana cellulare e può mantenere l'attività biologica. La forma precursore di IL-1 beta, a differenza del precursore di IL-1 alfa, mostra poca o nessuna attività biologica rispetto alla forma trasformata. Sia le forme non trasformate che quelle mature di IL-1 beta vengono esportate dalla cellula.
IL-1 alfa e IL-1 beta esercitano i loro effetti attraverso i recettori della superfamiglia delle immunoglobuline che legano inoltre IL-1ra. Il recettore transmembrana di tipo I da 80 kDa (IL-1 RI) è espresso su cellule T, fibroblasti, cheratinociti, cellule endoteliali, cellule del rivestimento sinoviale, condrociti ed epatociti. Il recettore transmembrana di tipo II da 68 kDa (IL-1 RII) è espresso sulle cellule B, sui neutrofili e sulle cellule del midollo osseo. I due tipi di recettori IL-1 mostrano circa il 28% di omologia nei loro domini extracellulari, ma differiscono significativamente in quanto il recettore di tipo II ha un dominio citoplasmatico di soli 29 aa, mentre il recettore di tipo I ha un dominio citoplasmatico di 213 aa. IL-1 RII non sembra segnalare in risposta a IL-1 e può funzionare come un recettore esca che attenua la funzione di IL-1. La proteina accessoria del recettore IL-1 (IL-1 RAcP) si associa a IL-1 RI ed è necessaria per la trasduzione del segnale di IL-1 RI. IL-1ra è una molecola secreta che funziona come un inibitore competitivo di IL-1. Forme solubili sia di IL-1 RI che di IL-1 RII sono state rilevate nel plasma umano, nei fluidi sinoviali e nei mezzi condizionati di diverse linee cellulari umane. Inoltre, le proteine leganti IL-1 che assomigliano a IL-1 RII solubile sono codificate dai virus del vaiolo vaccino e del vaiolo bovino.
IL-1 alfa e IL-1 beta sono polipeptidi strutturalmente correlati che mostrano circa il 25% di omologia a livello di amminoacidi (aa). Entrambi sono sintetizzati come precursori da 31 kDa che vengono successivamente scissi in proteine mature da circa 17,5 kDa. La scissione del precursore beta di IL-1 da parte di Caspase-1/ICE è un passaggio fondamentale nella risposta infiammatoria. Né IL-1 alfa né IL-1 beta contengono un tipico peptide segnale idrofobico, ma l'evidenza suggerisce che questi fattori possono essere secreti attraverso vie non-classiche. Una porzione di IL-1 alfa non trasformata può essere presente sulla membrana cellulare e può mantenere l'attività biologica. La forma precursore di IL-1 beta, a differenza del precursore di IL-1 alfa, mostra poca o nessuna attività biologica rispetto alla forma trasformata. Sia le forme non trasformate che quelle mature di IL-1 beta vengono esportate dalla cellula.
IL-1 alfa e IL-1 beta esercitano i loro effetti attraverso i recettori della superfamiglia delle immunoglobuline che legano inoltre IL-1ra. Il recettore transmembrana di tipo I da 80 kDa (IL-1 RI) è espresso su cellule T, fibroblasti, cheratinociti, cellule endoteliali, cellule del rivestimento sinoviale, condrociti ed epatociti. Il recettore transmembrana di tipo II da 68 kDa (IL-1 RII) è espresso sulle cellule B, sui neutrofili e sulle cellule del midollo osseo. I due tipi di recettori IL-1 mostrano circa il 28% di omologia nei loro domini extracellulari, ma differiscono significativamente in quanto il recettore di tipo II ha un dominio citoplasmatico di soli 29 aa, mentre il recettore di tipo I ha un dominio citoplasmatico di 213 aa. IL-1 RII non sembra segnalare in risposta a IL-1 e può funzionare come un recettore esca che attenua la funzione di IL-1. La proteina accessoria del recettore IL-1 (IL-1 RAcP) si associa a IL-1 RI ed è necessaria per la trasduzione del segnale di IL-1 RI. IL-1ra è una molecola secreta che funziona come un inibitore competitivo di IL-1. Forme solubili sia di IL-1 RI che di IL-1 RII sono state rilevate nel plasma umano, nei fluidi sinoviali e nei mezzi condizionati di diverse linee cellulari umane. Inoltre, le proteine leganti IL-1 che assomigliano a IL-1 RII solubile sono codificate dai virus del vaiolo vaccino e del vaiolo bovino.
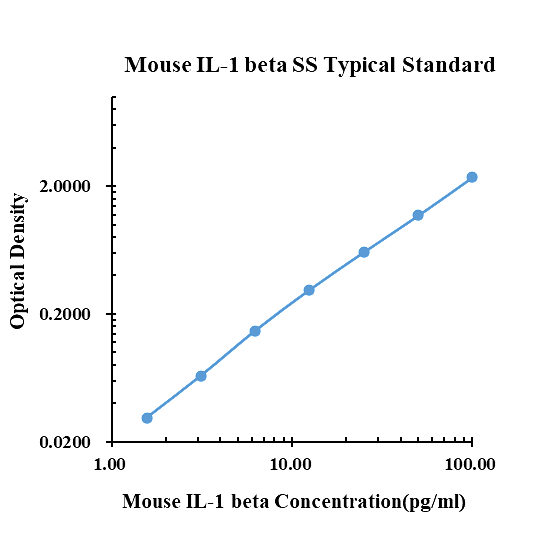
Dati tipici
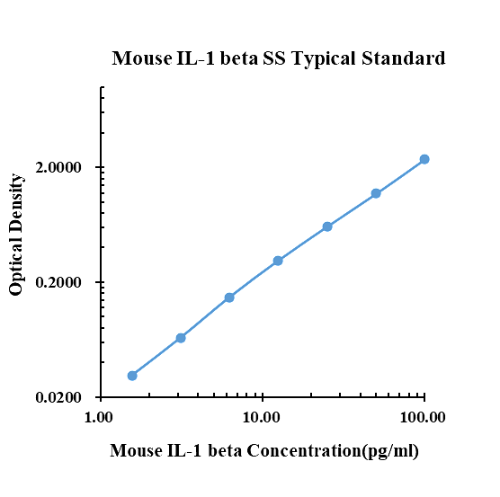
|
pg/ml |
D.O. |
Nella media |
Corretto |
|
|
0.00 |
0.1280 |
0.1360 |
0.1320 |
|
|
1.56 |
0.1530 |
0.1724 |
0.1627 |
0.0307 |
|
3.13 |
0.1902 |
0.2052 |
0.1977 |
0.0657 |
|
6.25 |
0.2792 |
0.2809 |
0.2801 |
0.1481 |
|
12.50 |
0.4370 |
0.4454 |
0.4412 |
0.3092 |
|
25.00 |
0.7625 |
0.7219 |
0.7422 |
0.6102 |
|
50.00 |
1.3050 |
1.3190 |
1.3120 |
1.1800 |
|
100.00 |
2.5070 |
2.4660 |
2.4865 |
2.3545 |
Precisione
|
Precisione intra-test |
Precisione inter-test |
|||||
|
Numero del campione |
S1 |
S2 |
S3 |
S1 |
S2 |
S3 |
|
22 |
22 |
22 |
6 |
6 |
6 |
|
|
Media (pg/ml) |
1.8 |
9.7 |
29.2 |
1.8 |
9.7 |
29.4 |
|
Deviazione standard |
0.030 |
0.005 |
0.002 |
0.032 |
0.005 |
0.002 |
|
Coefficiente di variazione(%) |
5.5 |
4.7 |
4.5 |
5.7 |
4.8 |
4.6 |
Recupero dei picchi
Il recupero del picco è stato valutato aggiungendo 3 livelli di IL-1 beta di topo a un campione di siero di topo sano. Il siero non addizionato è stato utilizzato come bianco in questo esperimento.
Il recupero variava dall'81% al 111% con un recupero medio complessivo del 98%.
Il recupero variava dall'81% al 111% con un recupero medio complessivo del 98%.
Valori campione
| Matrice del campione | Campione valutato | Intervallo (pg/ml) | Rilevabile (%) | Media rilevabile (pg/ml) |
|---|---|---|---|---|
| Siero | 30 | 0,18-3,93 | 100 | 1.47 |
Siero/Plasma – In questo test è stata valutata la presenza di IL-1 beta in trenta campioni provenienti da topi apparentemente sani. Non erano disponibili le cartelle cliniche dei donatori.
Nuovi prodotti