Kit ELISA IL-6 para ratas
 Folleto de protocolo
Folleto de protocolo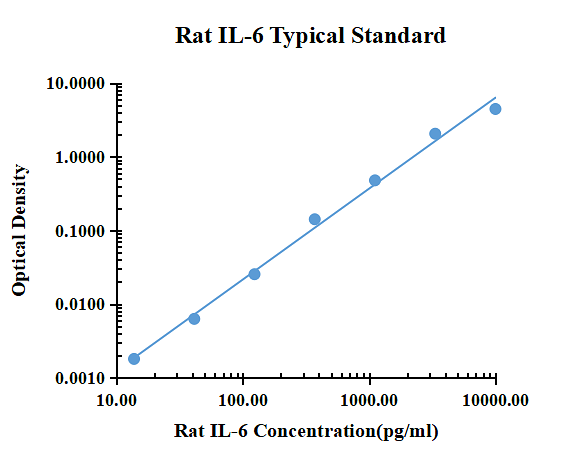
Características y detalles clave
- Especificación:
- Sensibilidad:
- Rango de curva estándar:
- Degradado de curva estándar:
- Número de incubaciones:
- Volumen de muestra:
- Tipo de ensayo:
- Duración de la operación:
-
Marca:
 Detalles del producto
Detalles del producto
IL-6 induce la señalización a través de un complejo receptor heterodimérico de la superficie celular compuesto por una subunidad de unión a ligando (IL-6 R alfa) y una subunidad transductora de señales (gp130). IL-6 se une a IL-6 Ra, lo que desencadena la asociación de IL-6 Ra con gp130 y la dimerización de gp130. gp130 también es un componente de los receptores para CLC, CNTF, CT-1, IL-11, IL-27, LIF y OSM. Las formas solubles de IL-6 Ra se generan mediante empalme alternativo y escisión proteolítica. En un mecanismo conocido como transseñalización, los complejos de IL-6 e IL-6 Ra solubles provocan respuestas de células que expresan gp130 que carecen de IL-6 Ra en la superficie celular. La transfirmación permite que una gama más amplia de tipos de células respondan a la IL-6, ya que la expresión de gp130 es ubicua, mientras que la de IL-6 Ra está predominantemente restringida a hepatocitos, monocitos y linfocitos en reposo. Las formas de empalme solubles de gp130 bloquean la transeñalización de IL-6/IL-6 Ra pero no de otras citocinas que utilizan gp130 como correceptor.
La IL-6, junto con el TNF-a y la IL-1, impulsa la respuesta inflamatoria aguda. La IL-6 es casi la única responsable de la fiebre y la respuesta de fase aguda en el hígado, y es importante en la transición de la inflamación aguda a la inmunidad adquirida o a la enfermedad inflamatoria crónica. Cuando está desregulado, contribuye a la inflamación crónica en afecciones como la obesidad, la resistencia a la insulina, la enfermedad inflamatoria intestinal, la artritis y la sepsis.
La IL-6 modula la resorción ósea y es un efector importante de la destrucción inflamatoria de las articulaciones en la artritis reumatoide a través de su promoción del desarrollo y la actividad de las células Th17. Contribuye al desarrollo y la desestabilización de la placa aterosclerótica, así como al desarrollo de la carcinogénesis asociada a la inflamación.
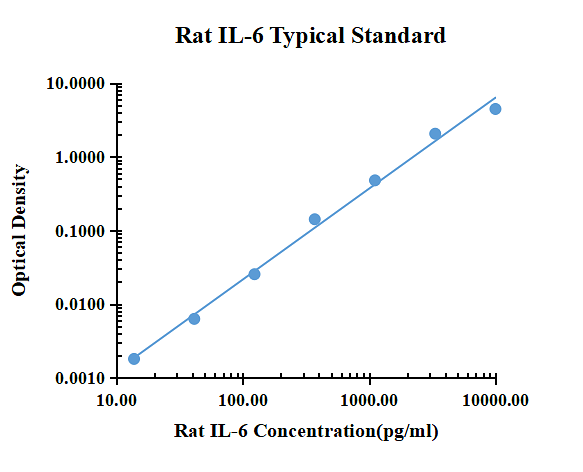
| pg/ml | D.O. | Promedio | Corregido | |
| 0.00 | 0.0579 | 0.0616 | 0.0598 | |
| 13.72 | 0.0653 | 0.0578 | 0.0616 | 0.0018 |
| 41.15 | 0.0688 | 0.0633 | 0.0661 | 0.0063 |
| 123.46 | 0.0853 | 0.0851 | 0.0852 | 0.0255 |
| 370.37 | 0.2024 | 0.2009 | 0.2017 | 0.1419 |
| 1111.11 | 0.5716 | 0.5064 | 0.5390 | 0.4793 |
| 3333.33 | 2.1332 | 2.1088 | 2.1210 | 2.0613 |
| 10000.00 | 4.5694 | 4.4840 | 4.5267 | 4.4670 |
| Precisión intra-ensayo | Precisión inter-ensayo | |||||
| Número de muestra | S1 | S2 | S3 | S1 | S2 | S3 |
| 22 | 22 | 22 | 6 | 6 | 6 | |
| Promedio (pg/ml) | 126.5 | 792.2 | 2890.8 | 155.2 | 987.5 | 3062.8 |
| Desviación estándar | 7.8 | 63.2 | 242 | 7.3 | 39.5 | 87.8 |
| Coeficiente de variación (%) | 6.2 | 8.0 | 8.4 | 4.7 | 4.0 | 2.9 |
Precisión intraensayo (Precisión dentro de un ensayo) Se analizaron tres muestras de concentración conocida veinte veces en una placa para evaluar la precisión intraensayo.
Precisión entre ensayos (Precisión entre ensayos) Se analizaron tres muestras de concentración conocida seis veces en una placa para evaluar la precisión intraensayo.
La recuperación del pico se evaluó añadiendo 3 niveles de IL-6 de rata a una muestra de suero de rata sana. El suero no enriquecido se utilizó como blanco en estos experimentos.
La recuperación osciló entre el 71% y el 99% con una recuperación media general del 89%.
| Matriz de muestra | Muestra evaluada | Rango (pg/ml) | Detectables (%) | Media de detectables (pg/ml) |
| suero | 30 | Dakota del Norte. | 0 | 0 |
Suero/plasma: se evaluaron treinta muestras de ratas aparentemente sanas para detectar la presencia de IL-6 en este ensayo. No se disponía de antecedentes médicos de los donantes.
Dakota del Norte. = no-detectable. Las muestras medidas por debajo de la sensibilidad se consideran no detectables.
Nuevos productos