Kit ELISA de súper sensibilidad IL-1β para ratón
Características y detalles clave
- Especificación:
- Sensibilidad:
- Rango de curva estándar:
- Degradado de curva estándar:
- Número de incubaciones:
- Volumen de muestra:
- Tipo de ensayo:
- Duración de la operación:
-
Marca:
CAT.NO. : AEMSS0029
US$ Por favor elige
US$ Por favor elige
Tamaño:
Sendero, tamaño a granel o solicitudes personalizadas Por favor contáctenos
 Detalles del producto
Detalles del producto
Antecedentes
La familia de proteínas Interleucina 1 (IL-1) está formada por IL-1 alfa, IL-1 beta y el antagonista del receptor de IL-1 (IL-1ra). La IL-1 alfa y la IL-1 beta se unen a los mismos receptores de la superficie celular y comparten funciones biológicas. La IL-1 no es producida por células no estimuladas de individuos sanos, a excepción de los queratinocitos de la piel, algunas células epiteliales y ciertas células del sistema nervioso central. Sin embargo, en respuesta a agentes inflamatorios, infecciones o endotoxinas microbianas, se observa un aumento espectacular en la producción de IL-1 por macrófagos y otros tipos de células. IL-1 beta desempeña un papel central en las respuestas inmunes e inflamatorias, la remodelación ósea, la fiebre, el metabolismo de los carbohidratos y la fisiología de GH/IGF-I. La producción inapropiada o prolongada de IL-1 se ha implicado en una variedad de condiciones patológicas que incluyen sepsis, artritis reumatoide, enfermedad inflamatoria intestinal, leucemia mielógena aguda y crónica, diabetes mellitus insulinodependiente, aterosclerosis, lesión neuronal y enfermedades relacionadas con el envejecimiento.
IL-1 alfa e IL-1 beta son polipéptidos estructuralmente relacionados que muestran aproximadamente un 25 % de homología a nivel de aminoácidos (aa). Ambos se sintetizan como precursores de 31 kDa que posteriormente se escinden en proteínas maduras de aproximadamente 17,5 kDa. La escisión del precursor IL-1 beta por Caspasa-1/ICE es un paso clave en la respuesta inflamatoria. Ni la IL-1 alfa ni la IL-1 beta contienen un péptido señal hidrofóbico típico, pero la evidencia sugiere que estos factores pueden secretarse por vías no clásicas. Una porción de IL-1 alfa sin procesar puede presentarse en la membrana celular y puede conservar actividad biológica. La forma precursora de IL-1 beta, a diferencia del precursor de IL-1 alfa, muestra poca o ninguna actividad biológica en comparación con la forma procesada. Tanto las formas maduras como las no procesadas de IL-1 beta se exportan desde la célula.
IL-1 alfa e IL-1 beta ejercen sus efectos a través de receptores de la superfamilia de inmunoglobulinas que además se unen a IL-1ra. El receptor transmembrana tipo I de 80 kDa (IL-1 RI) se expresa en células T, fibroblastos, queratinocitos, células endoteliales, células del revestimiento sinovial, condrocitos y hepatocitos. El receptor transmembrana tipo II de 68 kDa (IL-1 RII) se expresa en células B, neutrófilos y células de la médula ósea. Los dos tipos de receptores de IL-1 muestran aproximadamente 28% de homología en sus dominios extracelulares, pero difieren significativamente en que el receptor de tipo II tiene un dominio citoplasmático de sólo 29 aa, mientras que el receptor de tipo I tiene un dominio citoplasmático de 213 aa. IL-1 RII no parece emitir señales en respuesta a IL-1 y puede funcionar como un receptor señuelo que atenúa la función de IL-1. La proteína accesoria del receptor IL-1 (IL-1 RAcP) se asocia con IL-1 RI y es necesaria para la transducción de señales de IL-1 RI. IL-1ra es una molécula secretada que funciona como un inhibidor competitivo de IL-1. Se han detectado formas solubles de IL-1 RI e IL-1 RII en plasma humano, líquidos sinoviales y medios condicionados de varias líneas celulares humanas. Además, los virus vaccinia y de la viruela vacuna codifican proteínas de unión a IL-1 que se asemejan a la IL-1 RII soluble.
IL-1 alfa e IL-1 beta son polipéptidos estructuralmente relacionados que muestran aproximadamente un 25 % de homología a nivel de aminoácidos (aa). Ambos se sintetizan como precursores de 31 kDa que posteriormente se escinden en proteínas maduras de aproximadamente 17,5 kDa. La escisión del precursor IL-1 beta por Caspasa-1/ICE es un paso clave en la respuesta inflamatoria. Ni la IL-1 alfa ni la IL-1 beta contienen un péptido señal hidrofóbico típico, pero la evidencia sugiere que estos factores pueden secretarse por vías no clásicas. Una porción de IL-1 alfa sin procesar puede presentarse en la membrana celular y puede conservar actividad biológica. La forma precursora de IL-1 beta, a diferencia del precursor de IL-1 alfa, muestra poca o ninguna actividad biológica en comparación con la forma procesada. Tanto las formas maduras como las no procesadas de IL-1 beta se exportan desde la célula.
IL-1 alfa e IL-1 beta ejercen sus efectos a través de receptores de la superfamilia de inmunoglobulinas que además se unen a IL-1ra. El receptor transmembrana tipo I de 80 kDa (IL-1 RI) se expresa en células T, fibroblastos, queratinocitos, células endoteliales, células del revestimiento sinovial, condrocitos y hepatocitos. El receptor transmembrana tipo II de 68 kDa (IL-1 RII) se expresa en células B, neutrófilos y células de la médula ósea. Los dos tipos de receptores de IL-1 muestran aproximadamente 28% de homología en sus dominios extracelulares, pero difieren significativamente en que el receptor de tipo II tiene un dominio citoplasmático de sólo 29 aa, mientras que el receptor de tipo I tiene un dominio citoplasmático de 213 aa. IL-1 RII no parece emitir señales en respuesta a IL-1 y puede funcionar como un receptor señuelo que atenúa la función de IL-1. La proteína accesoria del receptor IL-1 (IL-1 RAcP) se asocia con IL-1 RI y es necesaria para la transducción de señales de IL-1 RI. IL-1ra es una molécula secretada que funciona como un inhibidor competitivo de IL-1. Se han detectado formas solubles de IL-1 RI e IL-1 RII en plasma humano, líquidos sinoviales y medios condicionados de varias líneas celulares humanas. Además, los virus vaccinia y de la viruela vacuna codifican proteínas de unión a IL-1 que se asemejan a la IL-1 RII soluble.
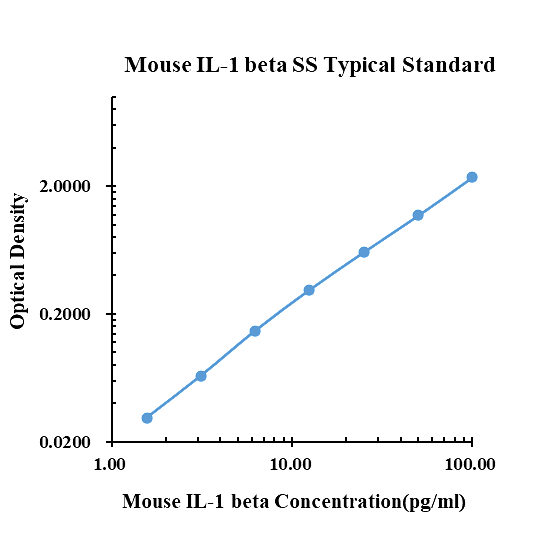
Datos típicos
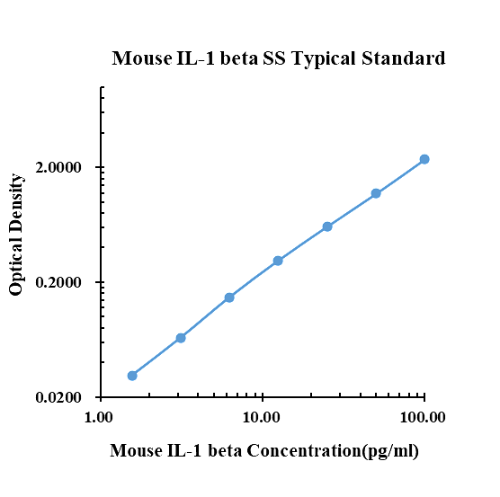
|
pg/ml |
D.O. |
Promedio |
Corregido |
|
|
0.00 |
0.1280 |
0.1360 |
0.1320 |
|
|
1.56 |
0.1530 |
0.1724 |
0.1627 |
0.0307 |
|
3.13 |
0.1902 |
0.2052 |
0.1977 |
0.0657 |
|
6.25 |
0.2792 |
0.2809 |
0.2801 |
0.1481 |
|
12.50 |
0.4370 |
0.4454 |
0.4412 |
0.3092 |
|
25.00 |
0.7625 |
0.7219 |
0.7422 |
0.6102 |
|
50.00 |
1.3050 |
1.3190 |
1.3120 |
1.1800 |
|
100.00 |
2.5070 |
2.4660 |
2.4865 |
2.3545 |
Precisión
|
Precisión intra-ensayo |
Precisión inter-ensayo |
|||||
|
Número de muestra |
S1 |
S2 |
S3 |
S1 |
S2 |
S3 |
|
22 |
22 |
22 |
6 |
6 |
6 |
|
|
Promedio (pg/ml) |
1.8 |
9.7 |
29.2 |
1.8 |
9.7 |
29.4 |
|
Desviación estándar |
0.030 |
0.005 |
0.002 |
0.032 |
0.005 |
0.002 |
|
Coeficiente de variación (%) |
5.5 |
4.7 |
4.5 |
5.7 |
4.8 |
4.6 |
Recuperación de picos
La recuperación del pico se evaluó añadiendo 3 niveles de IL-1 beta de ratón a una muestra de suero de ratón sano. El suero no enriquecido se utilizó como blanco en este experimento.
La recuperación osciló entre el 81% y el 111% con una recuperación media general del 98%.
La recuperación osciló entre el 81% y el 111% con una recuperación media general del 98%.
Valores de muestra
| Matriz de muestra | Muestra evaluada | Rango (pg/ml) | Detectables (%) | Media de detectables (pg/ml) |
|---|---|---|---|---|
| suero | 30 | 0,18-3,93 | 100 | 1.47 |
Suero/plasma: se evaluaron treinta muestras de ratones aparentemente sanos para detectar la presencia de IL-1 beta en este ensayo. No se disponía de antecedentes médicos de los donantes.
Nuevos productos